Problem 5: Rask oppdukking av arter i fossilregisteret støtter ikke darwinistisk evolusjon
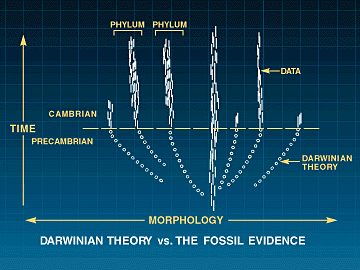 Fossil-registeret har lenge vært anerkjent som et problem for evolusjonsteorien. I 'Origin of Species' forklarte Darwin at teorien hans førte ham til å tro at "antallet av mellomvarianter, som tidligere har eksistert på jorden, virkelig må være enormt." (65) Han erkjente imidlertid at fossilene dokumenterte ikke disse 'mellomliggende' livsformene, og spurte: "Hvorfor er ikke hver geologisk formasjon og hvert fossilt lag fullt av slike mellomledd?" Darwins svar viste den spinkle karakteren til bevisene som støtter hans ideer: "Geologi avslører garantert ikke noen slik fint gradert organisk kjede; og dette er kanskje den mest åpenbare og alvorligste innvendingen som kan fremføres mot min teori." (67)
Fossil-registeret har lenge vært anerkjent som et problem for evolusjonsteorien. I 'Origin of Species' forklarte Darwin at teorien hans førte ham til å tro at "antallet av mellomvarianter, som tidligere har eksistert på jorden, virkelig må være enormt." (65) Han erkjente imidlertid at fossilene dokumenterte ikke disse 'mellomliggende' livsformene, og spurte: "Hvorfor er ikke hver geologisk formasjon og hvert fossilt lag fullt av slike mellomledd?" Darwins svar viste den spinkle karakteren til bevisene som støtter hans ideer: "Geologi avslører garantert ikke noen slik fint gradert organisk kjede; og dette er kanskje den mest åpenbare og alvorligste innvendingen som kan fremføres mot min teori." (67)
I dag, rundt 150 år senere, med tusenvis av arter kjent fra fossilregistrene, er det bare en liten brøkdel som hevdes å være kandidater for Darwins mellomformer. Fossile bevis på evolusjonære mellomprodukter mangler generelt, som den avdøde evolusjonære paleontologen Stephen Jay Gould innrømmet: "Fraværet av fossile bevis for mellomstadier mellom store overganger i organisk design, faktisk vår manglende evne, selv i vår fantasi, til å konstruere funksjonelle mellomprodukter i mange tilfeller, har vært et vedvarende og nagende problem for gradvise beretninger om evolusjon." (68)
Darwin forsøkte å redde sin teori om gradvis evolusjon ved å hevde at mellomliggende fossiler ikke blir funnet på grunn av "den ekstreme ufullkommenhet i den geologiske registreringen". (69) Selv Gould bemerket at Darwins argument om at fossilopptegnelsen er ufullkommen "vedvarer som den foretrukne flukt, av de fleste paleontologer, av forlegenhet for et register som ser ut til å vise så lite av direkte evolusjon." (70) Men i løpet av de siste tiårene har denne unnskyldningen mistet troverdighet.
 Paleontologer i dag erkjenner generelt at selv om fossil-registeret er ufullkomment, er det fortsatt tilstrekkelig til å vurdere spørsmål om evolusjon. En studie i Nature rapporterte at "hvis den skaleres til det … taksonomiske nivået til familien, gir de siste 540 millioner årene av fossilhistorien jevnt god dokumentasjon av fortidens liv." (71) En annen artikkel i Paleobiology evaluerte vår kunnskap om fossile registrere og konkluderte med at "vårt syn på historien til biologisk mangfold er modent." (72) Paleontologer erkjenner nå i økende grad at 'hopp' mellom arter, uten mellomformer , ikke bare er resultat av ufullstendig oversikt. Niles Eldredge, en evolusjonær paleontolog og kurator ved American Museum of Natural History, sier det slik med Ian Tattersal: "Registeret hopper, og alle bevis viser at registeret er ekte: hullene vi ser gjenspeiler virkelige hendelser i livets historie — ikke resultatet av et dårlig fossilregister." (73) Denne konklusjonen kom ikke lett, ettersom en vitenskapsmann som studerte under Gould følte behov for å bønnfalle kollegene sine om at "evolusjonsbiologer ikke lenger kan ignorere fossilregisteret på bakgrunn av at det er ufullkomment." (74)
Paleontologer i dag erkjenner generelt at selv om fossil-registeret er ufullkomment, er det fortsatt tilstrekkelig til å vurdere spørsmål om evolusjon. En studie i Nature rapporterte at "hvis den skaleres til det … taksonomiske nivået til familien, gir de siste 540 millioner årene av fossilhistorien jevnt god dokumentasjon av fortidens liv." (71) En annen artikkel i Paleobiology evaluerte vår kunnskap om fossile registrere og konkluderte med at "vårt syn på historien til biologisk mangfold er modent." (72) Paleontologer erkjenner nå i økende grad at 'hopp' mellom arter, uten mellomformer , ikke bare er resultat av ufullstendig oversikt. Niles Eldredge, en evolusjonær paleontolog og kurator ved American Museum of Natural History, sier det slik med Ian Tattersal: "Registeret hopper, og alle bevis viser at registeret er ekte: hullene vi ser gjenspeiler virkelige hendelser i livets historie — ikke resultatet av et dårlig fossilregister." (73) Denne konklusjonen kom ikke lett, ettersom en vitenskapsmann som studerte under Gould følte behov for å bønnfalle kollegene sine om at "evolusjonsbiologer ikke lenger kan ignorere fossilregisteret på bakgrunn av at det er ufullkomment." (74)
Et mønster av eksplosjoner
 Den endelige erkjennelsen av at fossilregistreringen ikke er helt ufullstendig, har tvunget evolusjonsbiologer til å akseptere at registreringen viser et mønster av eksplosjoner, ikke gradvis utvikling av levende organismer. En lærebok i biologi forklarer dette:
Den endelige erkjennelsen av at fossilregistreringen ikke er helt ufullstendig, har tvunget evolusjonsbiologer til å akseptere at registreringen viser et mønster av eksplosjoner, ikke gradvis utvikling av levende organismer. En lærebok i biologi forklarer dette:
Mange arter forblir praktisk talt uendret i millioner av år, for så å plutselig forsvinne for å bli erstattet av en ganske annen, men beslektet form. Dessuten dukker de fleste store dyregruppene brått opp i fossilregisteret, fullt utformet og uten fossiler ennå oppdaget, som danner en overgang fra foreldregruppen. (75)
Sannsynligvis er den mest kjente forekomsten av brå opptredener den kambriske eksplosjonen, hvor nesten alle de store levende dyrefylaene dukker opp i den kambriske perioden. En lærebok i biologi om virvelløse dyr forklarer dette:
De fleste av dyregruppene som er representert i fossilregisteret dukker først opp, 'fullt formet' og identifiserbare med hensyn til deres fylum, i Kambrium, for rundt 550 millioner år siden. Disse inkluderer så anatomisk komplekse og særegne typer som trilobitter, pigghuder, brachiopoder, bløtdyr og chordater. … Fossil-registeret er derfor ikke til hjelp med hensyn til opprinnelsen og tidlig diversifisering av de forskjellige dyrefylaene… (76)
 Evolusjonsforskere erkjenner at de ikke kan forklare denne raske opptredenen av forskjellige dyrekroppsplaner ved hjelp av klassiske darwinistiske prosesser, eller andre kjente materielle mekanismer. Robert Carroll, en paleontolog ved McGill University, hevder i Trends in Ecology and Evolution at "Den ekstreme hastigheten til anatomisk endring og adaptiv stråling i løpet av denne korte tidsperioden, krever forklaringer som går utover de som er foreslått for utviklingen av arter i den moderne biota." (77) En annen artikkel hevder likeledes at "mikroevolusjon ikke gir en tilfredsstillende forklaring på det ekstraordinære utbruddet av nyheter under den kambriske eksplosjonen" og konkluderer med at "de store evolusjonære overgangene i dyreevolusjon gjenstår fortsatt å være kausalt forklart." (78) Likeledes en artikkel fra 2009 i BioEssays innrømmer at "å belyse det materialistiske grunnlaget for den kambriske eksplosjonen har blitt mer unnvikende, ikke mindre, jo mer vi vet om selve hendelsen." (79)
Evolusjonsforskere erkjenner at de ikke kan forklare denne raske opptredenen av forskjellige dyrekroppsplaner ved hjelp av klassiske darwinistiske prosesser, eller andre kjente materielle mekanismer. Robert Carroll, en paleontolog ved McGill University, hevder i Trends in Ecology and Evolution at "Den ekstreme hastigheten til anatomisk endring og adaptiv stråling i løpet av denne korte tidsperioden, krever forklaringer som går utover de som er foreslått for utviklingen av arter i den moderne biota." (77) En annen artikkel hevder likeledes at "mikroevolusjon ikke gir en tilfredsstillende forklaring på det ekstraordinære utbruddet av nyheter under den kambriske eksplosjonen" og konkluderer med at "de store evolusjonære overgangene i dyreevolusjon gjenstår fortsatt å være kausalt forklart." (78) Likeledes en artikkel fra 2009 i BioEssays innrømmer at "å belyse det materialistiske grunnlaget for den kambriske eksplosjonen har blitt mer unnvikende, ikke mindre, jo mer vi vet om selve hendelsen." (79)
Men den kambriske eksplosjonen er på ingen måte den eneste eksplosjonen av liv som er registrert i fossilregisteret. Når det gjelder opprinnelsen til store fiskegrupper, skriver tidligere geoforsker Arthur Strahler fra Columbia University at "Dette er en telling i kreasjonistenes anklage som bare kan fremkalle unisont fra paleontologer en bønn om nolo contendere [ingen konkurranse]."80 En artikkel i Annual Review of Ecology and Systematics forklarer at opprinnelsen til landplanter "er den terrestriske ekvivalenten til den mye omdiskuterte kambriske 'eksplosjonen' av marine faunaer." (81)
 Når det gjelder opprinnelsen til angiospermer (blomstrende planter), har paleontologer oppdaget en 'stor blomstrings' -type eksplosjonshendelse. Som en artikkel sier:
Når det gjelder opprinnelsen til angiospermer (blomstrende planter), har paleontologer oppdaget en 'stor blomstrings' -type eksplosjonshendelse. Som en artikkel sier:
Til tross for mye forskning og analyser av forskjellige datakilder (f.eks. fossilregistrering og fylogenetiske analyser, ved bruk av molekylære og morfologiske karakterer), er opprinnelsen til angiospermene fortsatt uklar. Angiospermer dukker opp ganske plutselig i fossilregisteret ... uten noen åpenbare forfedre i en periode på 80-90 millioner år, før de dukket opp. (82)
På lignende måte opptrer mange pattedyrordener på en eksplosiv måte. Niles Eldredge forklarer at "det er alle slags hull: fravær av gradvis mellomliggende 'overgangsformer' mellom arter, men også mellom større grupper - mellom for eksempel familier av rovdyr, eller ordener av pattedyr." (83) Det er også en fugle-eksplosjon, med store fuglegrupper som dukket opp i løpet av en kort tidsperiode. (84) En artikkel i Trends in Ecology and Evolution med tittelen "Evolutionary Explosions and the Phylogenetic Fuse" forklarer:
-En bokstavelig lesning av fossilhistorien indikerer at tidlig kambrium (for ca. 545 millioner år siden) og tidlig tertiær (ca. 65 millioner år siden) var preget av enormt akselererte perioder med morfologisk evolusjon som markerte utseendet til dyrefylaene, og moderne henholdsvis fugle- og placenta-pattedyrordener. (85)
Selvfølgelig er det en håndfull eksempler der evolusjonsforskere tror de har funnet overgangsfossiler som dokumenterer gradvis darwinistisk evolusjon. Hvalens opprinnelse har blitt kalt en "reklame for makroevolusjon", (86) der det antas at for rundt 55 millioner år siden den mistet visse landpattedyr baklemmene og utviklet seg til helt vannlevende hvaler. Spesielt hevdes det at det er fossile landpattedyr med ørebein som ligner på hvaler, og fossile hvallignende pattedyr som beholder baklemmene.
Selv om virveldyr- og hvalekspert Phillip Gingerich innrømmer at vi bare har "fossiler som illustrerer tre eller fire trinn som bygger bro mellom hvalenes forløper til dagens pattedyr" (87), la oss for et øyeblikk anta at det eksisterer en full sekvens av fossiler. Er dette nok til å vise at denne overgangen skjedde? Selv om det er fossiler som ser ut som potensielle mellomformer, hvis den generelle evolusjonshistorien ikke gir mening, kan ikke fossilene være overgangsformer. I dette tilfellet står den darwinistiske utviklingen av hvaler fra landpattedyr overfor alvorlige matematiske utfordringer fra populasjonsgenetikk.
 Mange endringer ville vært nødvendige for å konvertere et landpattedyr til en hval, inkludert:
Mange endringer ville vært nødvendige for å konvertere et landpattedyr til en hval, inkludert:
Fremkomst av et blåsehull, med muskulatur og nervekontroll
Modifikasjon av øyet for permanent undervannssyn
Evne til å drikke sjøvann
Forlemmer forvandlet til svømmeføtter
Modifikasjon av skjelettstruktur
Evne til å amme unge under vann
Opprinnelsen til halefinner og muskulatur
Spekk for temperaturisolering (88)
Mange av disse nødvendige tilpasningene vil kreve flere koordinerte endringer. Men som vi så i Problem 3, krever slike samtidige mutasjoner ekstremt lange tidsperioder for å oppstå via den darwinistiske mekanismen. Hvalens utvikling går nå inn i et alvorlig problem. Fossilprotokollen krever at utviklingen av hvaler fra små landpattedyr måtte ha funnet sted på mindre enn 10 millioner år. (89) Det kan høres ut som lang tid, men det faller faktisk dramatisk kortvarig ut, spesielt gitt at hvaler har små bestandsstørrelser og lange generasjonstider. (90) Biolog Richard Sternberg har undersøkt kravene til denne overgangen matematisk og sier det slik: "For mange genetiske omkoblinger, for liten tid". (91)
Hvalens opprinnelse gir dermed en interessant case-studie av evolusjonære overganger: En sjelden gang der det faktisk finnes fossiler som potensielt viser mellomliggende egenskaper, blir ikke-styrt neo-darwinistisk evolusjon ugyldiggjort av den korte tiden som fossiler tillater. Hvis dette 'reklamen' for makroevolusjon ikke tåler gransking, hva forteller de oss om andre tilfeller, der evolusjonister viser frem antatte overgangsfossiler?
 Menneskelig opprinnelse og fossilregisteret
Menneskelig opprinnelse og fossilregisteret
Offentligheten blir faktisk ofte fortalt at det er fossiler som dokumenterer utviklingen av mennesker fra ape-lignende forløpere, men en nærmere titt på den tekniske litteraturen forteller en annen historie. Hominidfossiler faller generelt inn i en av to grupper: ape-lignende arter og menneskelignende arter, med et stort, uoverbygd gap mellom dem. I 2004 anerkjente den berømte evolusjonsbiologen Ernst Mayr menneskets brå opptreden:
-De tidligste fossilene av Homo, Homo rudolfensis og Homo erectus, er atskilt fra Australopithecus med et stort, uoverbygd gap. Hvordan kan vi forklare denne tilsynelatende vendingen? Uten å ha noen fossiler som kan tjene som 'missing links', må vi falle tilbake på den ærefulle metoden for historisk vitenskap, konstruksjonen av en historisk fortelling. (92)
I lys av slike bevis kalte en artikkel i Journal of Molecular Biology and Evolution utseendet til Homo sapiens "en genetisk revolusjon" der "ingen australopithecin-arter åpenbart er overgangsform." (93) Mangelen på fossile bevis for denne hypoteseovergangen, bekreftes av Harvard paleoantropologer Daniel E. Lieberman, David R. Pilbeam og Richard W. Wrangham:
-Av de forskjellige overgangene som skjedde under menneskelig evolusjon, var overgangen fra  Australopithecus til Homo utvilsomt en av de mest kritiske i sin omfang og konsekvenser. Som med mange viktige evolusjonære hendelser, er det både gode og dårlige nyheter. For det første er den dårlige nyheten at mange detaljer om denne overgangen er uklare på grunn av mangelen på fossile og arkeologiske registreringer. (94)
Australopithecus til Homo utvilsomt en av de mest kritiske i sin omfang og konsekvenser. Som med mange viktige evolusjonære hendelser, er det både gode og dårlige nyheter. For det første er den dårlige nyheten at mange detaljer om denne overgangen er uklare på grunn av mangelen på fossile og arkeologiske registreringer. (94)
Når det gjelder de "gode nyhetene", innrømmer de fortsatt: "selv om vi mangler mange detaljer om nøyaktig hvordan, når og hvor overgangen skjedde fra Australopithecus til Homo, har vi tilstrekkelige data fra før og etter overgangen til å trekke noen slutninger om overordnet karakter av nøkkelendringer som skjedde." (95) Med andre ord, fossiler gir ape-lignende australopithecines ('før'), og menneskelignende Homo ('etter'), men ikke fossiler som dokumenterer en overgang mellom dem. I fravær av mellomarter sitter vi igjen med 'slutninger' om en overgang kun basert på antagelsen om darwinistisk evolusjon. En kommentator foreslo at bevisene innebærer en 'big bang-teori' om utseendet til vår slekt Homo. (96) Dette gir ikke en overbevisende evolusjonær beretning om menneskelig opprinnelse. (97)
I stedet for å vise gradvis darwinistisk evolusjon, viser livets historien et mønster av eksplosjoner, der nye fossile former oppstår uten klare evolusjonære forløpere. Evolusjonsantropolog Jeffrey Schwartz oppsummerer problemet:
-Vi er fortsatt i mørket om opprinnelsen til de fleste større grupper av organismer. De vises i fossilregisteret slik Athena gjorde i hodet til Zevs - full ferdig og klar til å gå, i motsetning til Darwins skildring av evolusjon, som et resultat av den gradvise akkumuleringen av utallige, uendelig små variasjoner. . .” (98)
Dette utgjør en stor utfordring for darwinistisk evolusjon, inkludert synet på at alle dyr er i slekt gjennom felles avstamning.
Problem 6: Molekylærbiologi har ikke klart å gi et stort "livets tre"
 Da fossiler ikke klarte å demonstrere at dyr utviklet seg fra en felles stamfar, vendte evolusjonsforskere seg til en annen type bevis - DNA-sekvensdata - for å demonstrere et livets tre. På 1960-tallet, rundt den tiden den genetiske koden først ble forstått, antok biokjemikerne Émile Zuckerkandl og Linus Pauling at hvis DNA-sekvenser kunne brukes til å produsere evolusjonære trær - trær som matchet de som var basert på morfologiske eller anatomiske egenskaper - ville dette gi "det beste tilgjengelig enkeltbevis på makroevolusjonens virkelighet." (99) Dermed startet en tiår lang innsats for å sekvensere genene til mange organismer og konstruere 'molekylært' baserte evolusjonære ('fylogenetiske') trær. Det endelige målet har vært å konstruere et stort 'livets tre', som viser hvordan alle levende organismer er relatert gjennom universelle felles aner.
Da fossiler ikke klarte å demonstrere at dyr utviklet seg fra en felles stamfar, vendte evolusjonsforskere seg til en annen type bevis - DNA-sekvensdata - for å demonstrere et livets tre. På 1960-tallet, rundt den tiden den genetiske koden først ble forstått, antok biokjemikerne Émile Zuckerkandl og Linus Pauling at hvis DNA-sekvenser kunne brukes til å produsere evolusjonære trær - trær som matchet de som var basert på morfologiske eller anatomiske egenskaper - ville dette gi "det beste tilgjengelig enkeltbevis på makroevolusjonens virkelighet." (99) Dermed startet en tiår lang innsats for å sekvensere genene til mange organismer og konstruere 'molekylært' baserte evolusjonære ('fylogenetiske') trær. Det endelige målet har vært å konstruere et stort 'livets tre', som viser hvordan alle levende organismer er relatert gjennom universelle felles aner.
Hovedantakelsen
Den grunnleggende logikken bak bygging av molekylære trær er relativt enkel. Først velger etterforskerne et gen, eller en serie med gener, funnet på tvers av flere organismer. Deretter blir disse genene analysert for å bestemme deres nukleotidsekvenser, slik at gensekvensene til forskjellige organismer kan sammenlignes. Til slutt konstrueres et evolusjonstre basert på prinsippet om at jo mer lik nukleotidsekvensene er, desto nærmere beslektet arter. En artikkel i tidsskriftet Biological Theory sier det slik:
Molekulær systematikk er (stort sett) basert på antakelsen, først klart artikulert av Zuckerkandl og Pauling (1962), at graden av generell likhet reflekterer graden av slektskap. (100)
Denne antagelsen er i hovedsak en artikulering av et hovedtrekk ved teorien - ideen om universelt felles aner. Ikke desto mindre er det viktig å innse at det er en ren antagelse å hevde at genetiske likheter mellom ulike arter, nødvendigvis er et resultat av felles aner.
Om man opererer strengt innenfor et darwinistisk paradigme, følger disse antakelsene naturlig. Som den nevnte (Biological theory)-artikkelen forklarer, stammer hovedantagelsen som ligger til grunn for molekylære trær "fra å tolke molekylær likhet (eller ulikhet) mellom taxa i sammenheng med en darwinistisk modell for kontinuerlig og gradvis endring." (101) Så teorien antas å være sann til å bygge et tre. Men også, hvis darwinistisk evolusjon er sann, bør konstruksjon av trær ved hjelp av forskjellige sekvenser avsløre et rimelig konsistent mønster på tvers av forskjellige gener eller sekvenser.
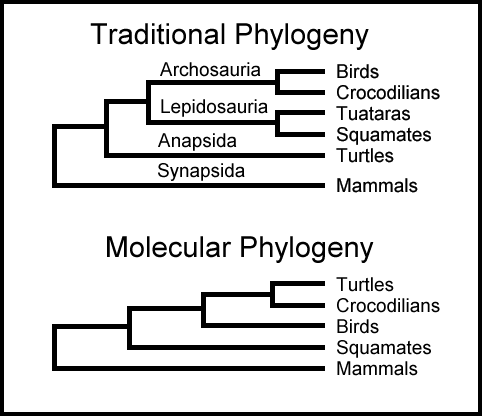 Dette gjør det desto mer betydningsfullt at forsøk på å bygge et storslått 'livets tre' ved å bruke DNA eller andre biologiske sekvensdata ikke har vært i samsvar med forventningene. Det grunnleggende problemet er at ett gen gir en versjon av livets tre, mens et annet gen gir en svært annerledes og motstridende versjon av treet. For eksempel, som vi vil diskutere videre nedenfor, plasserer standard pattedyrtreet, mennesker nærmere knyttet til gnagere enn til elefanter. Men studier av en viss type DNA kalt mikroRNA-gener har antydet det motsatte - at mennesker var nærmere elefanter enn gnagere. Slike konflikter mellom genbaserte trær er ekstremt vanlige.
Dette gjør det desto mer betydningsfullt at forsøk på å bygge et storslått 'livets tre' ved å bruke DNA eller andre biologiske sekvensdata ikke har vært i samsvar med forventningene. Det grunnleggende problemet er at ett gen gir en versjon av livets tre, mens et annet gen gir en svært annerledes og motstridende versjon av treet. For eksempel, som vi vil diskutere videre nedenfor, plasserer standard pattedyrtreet, mennesker nærmere knyttet til gnagere enn til elefanter. Men studier av en viss type DNA kalt mikroRNA-gener har antydet det motsatte - at mennesker var nærmere elefanter enn gnagere. Slike konflikter mellom genbaserte trær er ekstremt vanlige.
De genetiske dataene tegner dermed ikke et konsistent bilde av felles aner, og viser at antakelsene bak trebygging ofte feiler. Dette fører til berettigede spørsmål om universell felles avstamning er riktig.
Konflikter i bunnen av livets tre
Problemer oppsto først da molekylærbiologer sekvenserte gener fra de tre grunnleggende livsdomenene - bakterier, archaea og eukarya - men disse genene tillot ikke disse grunnleggende livsgruppene å bli knyttet til et trelignende mønster. I 2009 publiserte tidsskriftet New Scientist en forsidehistorie med tittelen "Hvorfor Darwin tok feil om livets tre" som forklarte disse dilemmaene:
-Problemene begynte tidlig på 1990-tallet da det ble mulig å sekvensere faktiske bakterielle og arkeale gener i stedet for bare RNA. Alle forventet at disse DNA-sekvensene skulle bekrefte RNA-treet, og noen ganger gjorde de det, men, avgjørende, noen ganger gjorde de det ikke. RNA, for eksempel, kan tyde på at art A var nærmere beslektet med art B enn art C, men et tre laget av DNA antyder det motsatte. (102)
Denne typen data fikk biokjemikeren W. Ford Doolittle til å forklare at "Molekylære fylogenister vil ikke ha klart å finne det 'sanne treet', ikke fordi metodene deres er utilstrekkelige eller fordi de har valgt feil gener, men fordi livets historie ikke ordentlig kan bli representert som et tre." (103) New Scientist sa det slik: "I lang tid skulle den hellige gral bygge et livstre … Men i dag ligger prosjektet i filler, revet i stykker av et angrep av negative bevis". (104)
Mange evolusjonister svarer noen ganger at disse problemene bare oppstår når man studerer mikroorganismer som bakterier - organismer som kan bytte gener gjennom en prosess som kalles "horisontal genoverføring", og dermed forkludre signalet om evolusjonære forhold. Men denne innvendingen er ikke helt sann, siden livets tre er utfordret selv blant høyere organismer, der slik genbytting ikke er utbredt. Carl Woese, en pioner innen evolusjonær molekylær systematikk, forklarer:
-Fylogenetiske uoverensstemmelser kan sees overalt i det universelle treet, fra roten til de store forgreningene innenfor og blant de forskjellige taxaene til sammensetningen av selve primærgruppene. (105)
 På samme måte bemerker New Scientist-artikkelen at "forskning tyder på at utviklingen av dyr og planter ikke er akkurat trelignende heller." (106) Artikkelen forklarer hva som skjedde da mikrobiolog Michael Syvanen prøvde å lage et tre som viser evolusjonære forhold ved å bruke 2000 gener fra en mangfoldig gruppe dyr:
På samme måte bemerker New Scientist-artikkelen at "forskning tyder på at utviklingen av dyr og planter ikke er akkurat trelignende heller." (106) Artikkelen forklarer hva som skjedde da mikrobiolog Michael Syvanen prøvde å lage et tre som viser evolusjonære forhold ved å bruke 2000 gener fra en mangfoldig gruppe dyr:
Han feilet. Problemet var at ulike gener fortalte motstridende evolusjonære historier. ... genene sendte blandede signaler. … Omtrent 50 prosent av genene har én evolusjonshistorie og 50 prosent en annen. (107)
Dataene var så vanskelige å løse opp i et tre at Syvanen beklaget: "Vi har nettopp utslettet livets tre." (108) Mange andre artikler i den tekniske litteraturen gjenkjenner lignende problemer.
Konflikter mellom høyere grener
En artikkel fra 2009 i Trends in Ecology and Evolution bemerker at "En stor utfordring for å inkorporere så store mengder data i slutninger om artstrær, er at motstridende slektshistorier ofte eksisterer i forskjellige gener gjennom genomet." (109) På samme måte, studerte en artikkel i Genome Forskning DNA-sekvensene i forskjellige dyregrupper og fant at "ulike proteiner genererer forskjellige fylogenetiske tre." (110) En artikkel i Nature i juni 2012 rapporterte at korte RNA-tråder kalt mikroRNA "er i ferd med å rive i stykker tradisjonelle ideer om dyrefamilien tre." Dartmouth-biolog Kevin Peterson, som studerer mikroRNA, beklaget: "Jeg har sett på tusenvis av mikroRNA-gener, og jeg kan ikke finne et eneste eksempel som støtter det tradisjonelle treet." I følge artikkelen ga mikroRNA "et radikalt annerledes diagram for pattedyr: et som justerer mennesker tettere med elefanter enn med gnagere." Peterson sa det rett ut: "MikroRNA-ene er helt entydige ... de gir et helt annet tre enn det alle andre ønsker." (111)
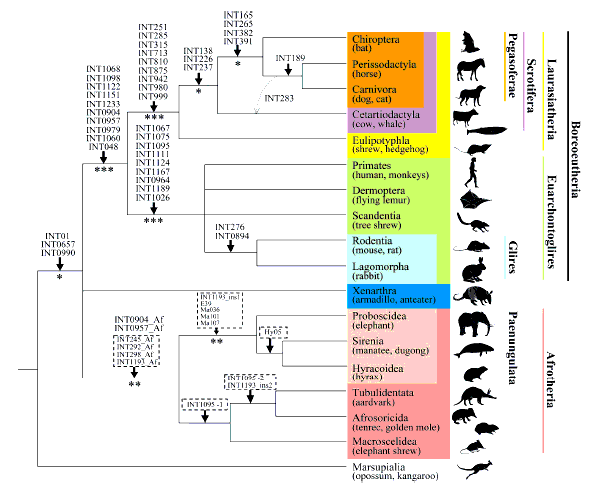 Konflikter mellom molekyler og morfologi
Konflikter mellom molekyler og morfologi
Ikke alle fylogenetiske trær er konstruert ved å sammenligne molekyler som DNA fra forskjellige arter. Mange trær er basert på å sammenligne formen, strukturen og kroppsplanen til forskjellige organismer - også kalt "morfologi." Men konflikter mellom molekylbaserte trær og morfologibaserte trær er også vanlig. En artikkel fra 2012 som studerte flaggermus-forhold gjorde dette klart, og sa: "Inkongruens mellom fylogenier avledet fra morfologiske versus molekylære analyser, og mellom trær basert på forskjellige undergrupper av molekylære sekvenser har blitt gjennomgripende ettersom datasett har utvidet seg raskt i både karakterer og arter." (112) Dette er neppe den eneste studien som møter konflikter mellom DNA-baserte trær og trær basert på anatomiske eller morfologiske egenskaper. Lærebøker hevder ofte at vanlig avstamning støttes ved å bruke eksemplet med et dyretre basert på enzymet Cytokrom C som samsvarer med det tradisjonelle evolusjonstreet basert på morfologi. (113) Lærebøker nevner imidlertid sjelden at treet basert på et annet enzym, Cytokrom B, kommer skarpt i konflikt med standard evolusjonstreet.
Som en artikkel i Trends in Ecology and Evolution observerte:
Det mitokondrielle Cytokrom B-genet antydet. . . en absurd fylogeni av pattedyr, uavhengig av metoden for trekonstruksjon. Katter og hvaler falt innenfor primater, gruppert med simianer (aper og gorillaer) og strepsirhiner (lemurer, bush-babyer og loriser) med unntak av tarsiere. Cytokrom B er sannsynligvis det mest sekvenserte genet hos virveldyr, noe som gjør dette overraskende resultatet enda mer foruroligende. (114)
Påfallende nok konkluderte en annen artikkel i Trends in Ecology and Evolution, "rikdommen av konkurrerende morfologiske, så vel som molekylære forslag av de rådende fylogeniene til pattedyrordene, ville redusere [pattedyrtreet] til et uløst buskas, den eneste konsistente busken. [evolusjonært forhold] er sannsynligvis grupperingen av elefanter og sjøkuer." (115) På grunn av slike konflikter, rapporterte en stor oversiktsartikkel i Nature, "forskjeller mellom molekylære og morfologiske trær" fører til 'evolusjonskrig' fordi evolusjonære trær konstruert ved å studere biologiske molekyler ligner ofte ikke på de som er trukket opp fra morfologi." (116)
 Til slutt prøvde en studie publisert i Science i 2005 å bruke gener for å rekonstruere relasjonene til dyrefylaene, men konkluderte med at "Til tross for mengden data og bredden av taxaene som ble analysert, forble forholdet mellom de fleste [dyre] phyla uavklarte. ” året etter publiserte de samme forfatterne en vitenskapelig artikkel med tittelen "Bushes in the Tree of Life", som ga slående konklusjoner. Forfatterne erkjenner at "en stor brøkdel av enkeltgener produserer fylogenier av dårlig kvalitet," og observerte at en studie "utelot 35 % av enkeltgener fra datamatrisen deres, fordi disse genene produserte fylogenier i strid med konvensjonell visdom." Artikkelen antyder at "visse kritiske deler av livets tre kan være vanskelig å løse, uavhengig av mengden konvensjonelle data som er tilgjengelig." Artikkelen hevder til og med at "den tilbakevendende oppdagelsen av vedvarende uløste klader (busker), bør tvinge frem en revurdering av flere utbredte antakelser om molekylær systematikk." (117)
Til slutt prøvde en studie publisert i Science i 2005 å bruke gener for å rekonstruere relasjonene til dyrefylaene, men konkluderte med at "Til tross for mengden data og bredden av taxaene som ble analysert, forble forholdet mellom de fleste [dyre] phyla uavklarte. ” året etter publiserte de samme forfatterne en vitenskapelig artikkel med tittelen "Bushes in the Tree of Life", som ga slående konklusjoner. Forfatterne erkjenner at "en stor brøkdel av enkeltgener produserer fylogenier av dårlig kvalitet," og observerte at en studie "utelot 35 % av enkeltgener fra datamatrisen deres, fordi disse genene produserte fylogenier i strid med konvensjonell visdom." Artikkelen antyder at "visse kritiske deler av livets tre kan være vanskelig å løse, uavhengig av mengden konvensjonelle data som er tilgjengelig." Artikkelen hevder til og med at "den tilbakevendende oppdagelsen av vedvarende uløste klader (busker), bør tvinge frem en revurdering av flere utbredte antakelser om molekylær systematikk." (117)
Dessverre er en antakelse om at disse evolusjonsbiologene ikke er villige til å revurdere, antagelsen om at universelle felles aner er korrekt. De appellerer til en myriade av ad hoc-argumenter – horisontal genoverføring, lang grenattraksjon, rask evolusjon, ulike utviklingshastigheter, koalescentteori, ufullstendig prøvetaking, feilaktig metodikk og konvergent evolusjon – for å bortforklare ubeleilig data som ikke passer til ettertraktet trelignende mønster. Som en artikkel fra 2012 uttalte, "fylogenetisk konflikt er vanlig, og ofte normen snarere enn unntaket." (118) På slutten av dagen har drømmen om at DNA-sekvensdata ville passe inn i et pent og pent livstre mislyktes, og med det en nøkkelprediksjon fra neo-darwinistisk teori.
Oversettelse og bilder ved Asbjørn E. Lund